
In the upper right corner is a “plate”: the microplates with 96 wells where we deposit small tissue samples that are to be processed at the CCDB lab in Canada for NorBOL
On the third day of Christmas,
we sent eleven microplates away:
one plate cnidarians (A)
two with worms a-wriggle (B)
two plates of insects (C)
three plates crustaceans (D)
two (and a half) plates of mites (E)
and a half-plate assorted a-arthropods (F)!
Ahem. Yes.
As Endre explained in the fifth post of the calendar, collecting, identifying, documenting and keeping specimens used for DNA barcoding is an important part of what we do here at the invertebrate collections. Our mission in the NORBOL consortium is to produce DNA-barcodes, particularly for marine fauna in Norwegian waters and to make these barcodes available with open access to records and metadata in the BOLD database. These samples contribute to the building of a validated reference library of the genetic barcodes of the species found in Norway. You can search for different taxonomic groups here to see if they have been barcoded from Norwegian territory: Search NorBOL
The process is fairly straight forward (at least on paper!): Animals are collected and identified. Those species relevant for barcoding are selected, and a specimen (=1 animal) is chosen to be barcoded. We take a small tissue sample from the specimen, and keep the rest of the animal as the barcode voucher; if the need should arise to check if it really is what we initially thought, it is crucial to be able to go back and check the animal again. The tissue samples are collected in wells on a plate like the one pictured above, and the information about the animals – where they were collected, who collected them, what species they are, who identified them and so on is uploaded to BOLD together with images of the animals.

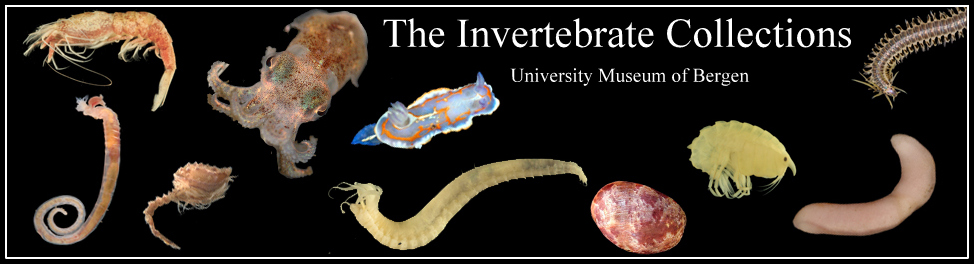
Representatives for the tissue sample plates that we just shipped off. Thank you Steffen, Anna and Per for contributing the terrestrial animals and images! Photos: L. Martell, A. Seniczak, S. Roth, K. Kongshavn. Illustration: K. Kongshavn
On Monday we shipped a new batch of plates – as (attempted) illustrated in song above.
Included is material from several of the Norwegian Taxonomy Initiative projects (artsprosjekt) that are happening at the University Museum of Bergen. We are coordinating the efforts on marine life, but are of course also facilitating the NorBOL barcoding of other organisms that take place at the UMB. There are animals from NorAmph (Norwegian Amphipoda), Hydrozoan pelagic diversity in Norway (HYPNO), Orbatid mites, and the insects found associated with nutrient rich marshes in Hedmark in this shipment.
We have also prepared several plates of Crustaceans collected and identified by the Norwegian marine mapping programme Mareano – one of the great contributors of material to the collections.
Now we wait for the lab to process them, and for the genetic sequences to be uploaded to BOLD – fingers crossed for many interesting results!
-Katrine