“Julebukken” – the Yule Goat.
A goat made of straw has commonly been observed among the paraphernalia that people put on display around Christmas times in the Nordic countries. Ask people what they symbolize and I bet that the majority would say just “Christmas” without having a further explanation at hand. It is very likely that the Yule Goat is a remnant from the pagan celebrations of the December solstice. The mythological origin of the Yule Goat is unclear (see Gunnell 1995). It is probably of mixed origin because cultural evolution is often syncretic, – a blend of beliefs, mythologies, and practices from different sources and “ethno-folkloristic schools of thought”. It has been speculated that the straw figure of the Yule Goat reflects some sort of pagan vegetation god ruling over grain growth and who required particular human attention around the winter solstice.

“Julebukk” – a common Christmas decoration.
Others have associated Yule Goat with the Germanic thunder-god Thor, because he used two bucks to pull his chariot over the sky. Thor must have been an environmentally friendly god because these bucks were used as a food resource in Valhalla and if only he took great care to keep the bones and the skin, he could revive fully reincarnated goats the next day. The idea of a resurrected goat was dramatized in folk rituals of Nordic small communities by people who reenacted the death and revival of the Yule Buck in songs and theater performed on house visits by a ragged assembly of masked people. The central character in these folkloristic plays was a person either wrapped in straw or hides and carrying a goat head, sometimes also a hammer (like Thor). Right up to the mid nineteenth century, plays like these were practiced in Scandinavia. The traditions have several characteristics in common with Halloween and the Yule Buck masquerade apparently is a tradition that now seems to be fading and to be replaced by Halloween celebrations. In both cases there is an underlying theme of temporary breakdown and restoration of cosmological and moral order when cyclic time has gone the full circle. A small scale version of this idea is also at work around midnight, – the ghost hour. The Cristian tradition has tended to associate goats with naughty behaviours of all sorts, particularly in terms of sexuality. Goats were also at times associated with mythological creatures like Pan and with the Devil.
The Star Buck
The Babylonians divided the sky in 360 parts, but ancient astronomers also used twelve 30 degrees sectors of the sky to reference the positions of celestial bodies on the ecliptic, – the track that the sun appears to follow through the year. Like the classic analogous clock with twelve sectors marking divisions of the day and night the zodiac is a clock for the earth’s revolution around the sun. Because the rotation axis of the earth is tilted, the sun appears to draw an S-shaped path around the Earth. From a northern perspective the maximum of the path is the summer solstice. The minimum is the winter solstice, when the zenith of the sun is farthest away from the Arctic and the days are shortest on the Northern hemisphere. The exact time for the solstice is not easy to determine, but ancient astronomers found that the winter solstice took place when the sun was in the sector of the star constellation called Capricornus. Claudius Ptolemy, who is known as the prime authority of pre-Copernican cosmological texts, wrote in Book 1 chapter 11 of his very influential Tetrabiblos :
“For the sun turns when he is at the beginning of these signs and reverses his latitudinal progress, causing summer in Cancer and winter in Capricorn.”
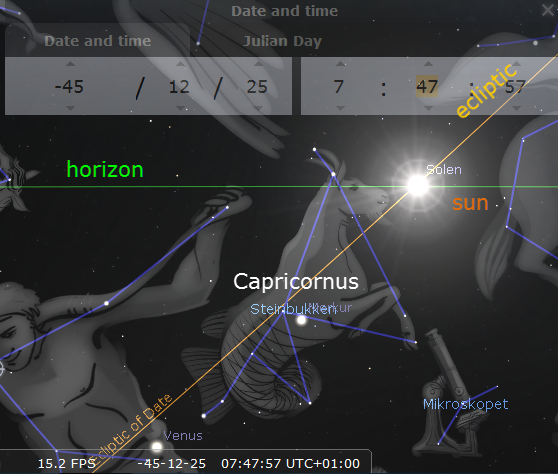
Capricornus is Latin for “goat horn” and Capricorn is sometimes depicted as a goat, sometimes as half goat, half fish. Because the “turning point” of the sun at winter solstice was once at an imaginary latitude circle drawn through Capricornus, we say that the sun is turning at the Tropic of Capricorn. However, due to the swaying of the Earth’s rotation axis, winter solstice is no longer in Capricorn and the Tropic of Capricorn has also moved away so that, paradoxically, the Tropic of Capricorn is now passing through Sagittarius . When the Julian calendar took effect 45 years BC the solstice was celebrated on 25th of December, but apparently winter solstice was already about to leave Capricornus in the direction of Sagittarius. Historians of astronomy think that Capricornus was already a marker of seasonal time about 2000 years BC.
The sun in Capricornus seen from Rome 45 years BP according to Stellarium.
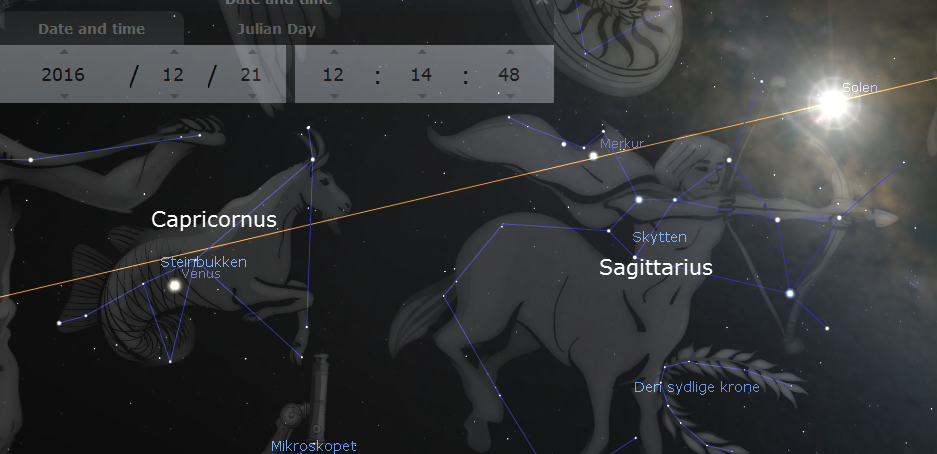
The sun at winter solstice in 2016 seen from Rome according to Stellarium.
Although much speculation has been put forward about the origins of Yule Buck, I suspect that the role of the goat in the sky has been undervalued when trying to understand the conducts and traditions of people in the Nordic countries around the winter solstice. Surely, the teaching of Ptolemy must have diffused somehow to ordinary people during the Medieval Ages. After all, Capricorn was the messenger of a better existence to come, if one could only sustain over the winter.
Resurrecting a goat
Goats that stood model for the Capricorn have been part of the human environments since long before they were painted on cave walls in Ardeche at the foot of the Pyrenees about 30000 years BC. Archaeological material from Jordan indicates that goats were domesticated already 7000 BC as one of the first of the ruminant species. Wild goats are members of the genus Capra and are distributed with several species over the Eurasian and African continents. Most of the wild goats are now regarded as more or less threatened species due to hunting pressure and habitat loss. In Spain the so-called bucardo goat was declared extinct in year 2000, when the last individual was hit by a falling tree in Oresa National Park.

The extinct subspecies of the Pyrenean Ibex. (Source Wikipedia)
An international group of biotechnologists set up experiments to resurrect the bucardo, which is considered as a subspecies of the Pyrenean Ibex, Capra pyrenaica. It is perhaps not likely that these scientists were inspired by the Nordic myth about Thor and his perpetual buck-goats. Nevertheless, they had already taken skin samples from the last living female the year before she died. With the frozen cells from the skin they had cellular nuclei with goat genome and also a plasma membrane with small amounts of cytoplasma. With a technique similar to the one that was pioneered by the Roslin Institute in Edinburgh to clone the sheep Dolly, they replaced the cell contents of domestic goat eggs with somatic cell material from the dead ibex. Then they implanted the manipulated eggs into many substitute mothers of both domesticated goats and of females of the Spanish ibex. Only one of the embryos survived long enough to be released by cesarean section and it died only a few minutes after birth. Despite this and similar failed efforts to reconstruct extinct evolutionary lineages, it is still claimed by scientists who are involved in this business that such methods hold promise for rescuing rare and endangered species. But without making claims of being a specialist in conservation genetics I ask myself: how is it possible to save a species when all of the genetic variability in the population is lost. This may have been a problem already before the population went extinct because the sets of genes that run the immune system and code for the proteins that protect the body from foreign molecules, the so-called major histocompatibility complex, had already been observed to lack variability. Then of course, in the case of the extinct Pyrenean ibex there is also another problem that would be a major impediment in the reconstruction of a natural population. It is a problem of the missing Y-chromosome. Goat Y-chromosomes would be necessary to have functional males of reconstructed goats. Basically this is a problem that has also puzzled thinkers with respect to the child that allegedly was born by a virgin on the solstice 2000 years ago. Did he have a chromosome set from the father?
Merry solstice, Christmas, and Happy New Year!
EW
More reading
Folch, J. et al. (2009) First birth of an animal from an extinct subspecies (Capra pyrenaica pyrenaica) by cloning. Theriogenology 71:1026–1034.
Gunnell, T. (1995) The Origins of Drama in Scandinavia. Boydell & Brewer Ltd.
Hinson, R. (2015) Goat. Reaction Books, London.
Lee, K. (2001) Can cloning save endangered species? Current Biology 11 (7): R245-R246